Our Species
Cactophilic Drosophila oviposit and feed as larvae and adults on the necrotic tissues of different species of cacti. These flies are largely obtaining nutrients from the yeasts and bacteria that reside in necrotic cacti, but also take up and metabolize cactus-derived compounds, some of which can be toxic to other species. In this system the host environment is produced by an interaction between the composition of the cactus tissues and the communities of yeasts and bacteria involved in their decomposition; different species of cactus have distinct microbial community offering distinct chemical profiles in the necroses.
Luciano Matzkin
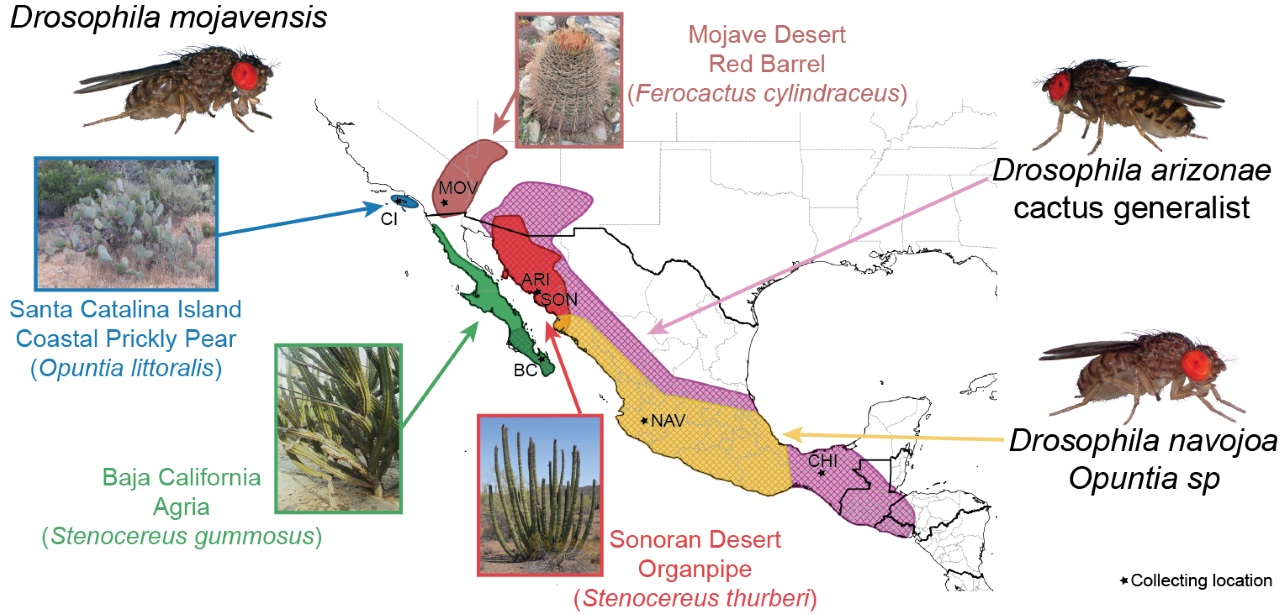
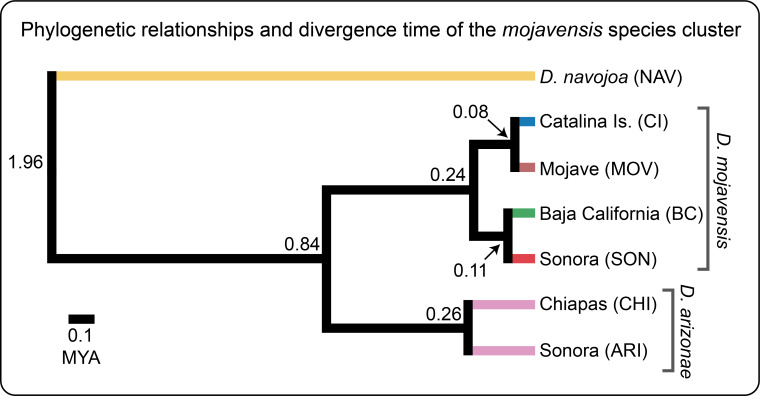
The mojavensis species cluster is nested within the larger repleta group of species. Within the mojavensis cluster, D. mojavensis and D. arizonae are sister species, with D. navojoa as a relatively more distant outgroup. All three are cactophilic, with D. navojoa being an Opuntia spp. breeder found in central Mexico, D. arizonae a cactus generalist mostly utilizing columnar species with a broad distribution from southern North America to the U.S., and D. mojavensis, composed of four genetically and ecologically divergent cactus host populations. The four D. mojavensis cactus host populations are: Catalina Island using coastal prickly pear (Opuntia littoralis), Mojave Desert using red barrel (Ferocactus cylindraceus), Baja California using agria (Stenocereus gummosus) and Sonora Desert using organpipe (S. thurberi). Our most recent phylogenomic analysis indicates a divergence time of ~0.84 MYA between the sister species and much less between the host populations. The close evolutionary relationships as well as the distinct local ecological conditions of each of these populations and species make it an excellent and powerful system to study the evolution of adaptive phenotypes and reproductive incompatibilities.
General selected references
Barker JSF, Starmer WT. 1982. Ecological genetics and evolution: the cactus-yeast-Drosophila model system. Sydney ; New York: Academic Press.
Fellows, D.F. & Heed, W.B. Factors affecting host plant selection in desert-adapted cactiphilic Drosophila. Ecology 53, 850-858 (1972).
Heed, W.B. Ecology and genetics of Sonoran desert Drosophila. in Ecological genetics: The interface (ed. Brussard, P.F.) 109-126 (Springer-Verlag, 1978).
Matzkin LM. 2014. Ecological Genomics of Host Shifts in Drosophila mojavensis. In: Landry C, Aubin-Horth N, editors. Ecological Genomics: Ecology and the Evolution of Genes and Genomes. New York: Springer. p. 223-247.
Ruiz A, Heed WB, Wasserman M. 1990. Evolution of the Mojavensis cluster of cactophilic Drosophila with descriptions of two new species. Journal of Heredity 81:30-42.
Starmer WT, Aberdeen V. 1990. The nutritional importance of pure and mixed cultures of yeasts in the development of Drosophila mulleri larvae in Opuntia tissues and its relationship to host plant shifts. In: Barker JSF, Starmer WT, MacIntyre RJ, editors. Ecological and evolutionary genetics of Drosophila. New York: Plenum Press. p. 145-160.
Genome line information
| Species | Population Abb. | Collection Location/Date | D. species Stock Center ID | Old ID |
|---|---|---|---|---|
| D. mojavensis | BC | La Paz, Baja California, Mexico (2001) | 15081-1354.01 | MJBC 155 |
| CI | Santa Catalina Island, California, US (2002) | 15081-1352.22 | 15081-1352.22 | |
| MOV | Anza-Borrego State Desert Park, California, US (2002) | 15081-1353.01 | MJANZA 402-8 | |
| SON | Guaymas, Sonora, Mexico (1998) | 15081-1355.01 | MJ 122 | |
| D. arizonae | ARI | Guaymas, Sonora, Mexico (2004) | 15081-1271.41 | AR002 |
| CHI | Chiapas, Mexico (1987) | 15081-1271.14 | AZ Chiapas 1B 13610 | |
| D. navojoa | NAV | Jalisco, Mexico (1997) | 15081-1374.11 | 15081-1374.11 |
Some lines may not be available currently from the Drosophila Species Stock Center contact us directly if interested in obtaining them.